

The Effect of Suppressing Discoidin Domain Receptor Expression on Keloid Formation and Proliferation
Abstract
Background. Discoidin domain receptors (DDR) with tyrosine kinase activity have been identified as novel receptors for modulating collagen production and organization in scar tissue. The purpose of this study was to explore the effect of blocking discoidin domain receptor 1 (DDR1), signaling of keloid fibroblast cells on the inhibition of keloid formation, and proliferation, by means of gene therapy. Methods. The expression of DDR1 in keloid tissues was investigated by immunohistology and the expression of DDR1 protein in keloid fibroblasts was examined by Western blot analysis. Keloid dermal fibroblasts were infected in vitro with modified phosphorothioate and liposome-encapsulation DDR1-antisense oligodeoxynucleotide (ASODN). Northern blot was used to analyze gene expression of DDR1 in infected keloid dermal fibroblasts and the effect on type I and type III collagen gene expression. Extracellular matrix production in infected fibroblasts was analyzed by [3H] proline incorporation. Results. In keloid tissues, the expression of DDR1 was observed to be widely and strongly distributed. The expression of DDR1 protein was also highly increased in keloid fibroblasts compared to normal skin fibroblasts. This was markedly downregulated in lipid-encoding DDR1-ASODN infected fibroblasts compared to lipid encoding DDR1-NSODN infected fibroblasts and lipid-infected fibroblasts. Type I and type III collagen gene expression and extracellular matrix production also were downregulated markedly in DDR1 ASODN infected fibroblasts. Moreover, the ratio of type I and type III collagen was significantly improved. Conclusion. An intrinsic functional difference exists between normal human dermal and keloid fibroblasts in terms of higher DDR1 gene expression in keloid fibroblasts. The quality and quantity of collagen can be improved by downregulating the expression of DDR1 using ASODN. This intervention is potentially useful in controlling fibrosis and keloid formation in clinical settings.
Introduction
Successful wound healing involves a complete orchestration of cytokines, growth factors, and extracellular matrix (ECM) components. The regulation of ECM components, especially collagen, is key in tissue remodeling and represents a vital step in the wound healing process, with or without scarring. Keloids are abnormal scars that grow beyond the boundary of the original site of skin injury and are characterized by excess accumulation of ECM components. The underlying mechanisms of pathogenesis remain unknown. Clinically, the keloid is a significant cosmetic problem, yet present treatment methods remain largely ineffective.1
Many hypotheses have attempted to explain the etiology of keloid formation,2 and many studies have been devoted to the comparison of normal dermal and keloid fibroblasts.3,14–16 Recently, researchers have focused on transforming growth factor-beta (TGF-β),3 which has a probable role in scar formation.
Additionally, cellular signaling mechanisms may indicate inherent differences in response to injury between keloid fibroblasts and normal dermal fibroblasts Specifically, the major difference in gene regulation that leads to abnormal excess production of ECM in keloid fibroblasts is due to the interaction of cell surface receptors and the ECM.
Wound healing is a dynamic biological process involving many cell-cell and cell-matrix interactions in a complex milieu. Keloids are a result of an “overhealed” wound. The interaction of ECM and the cell result in abnormal cell behavior. An essential factor exists in the signaling pathways, which exert their function by altering the phosphorylation state of tyrosine residues of the target protein in the cell membrane. A recent hypothesis suggested dysregulated-sensing mechanisms in keloid also might contribute to its formation. Discoidin domain receptors 1 and 2 (DDR1 and DDR2), which recently have been identified as collagen receptors, are thought to play important roles in sensing the differences of spatial and temporal distribution of collagen components.1,7
Functionally, DDR1 and DDR2 are tyrosine kinase receptors that are composed of a dictyostelium disodium protein discoidin in their extracellular region, transmembrane domain, and a highly conserved catalytic domain.8 Collagen directly bound to the receptor ectodomain results in phosphorylation of the tyrosine kinase domain, which in turn leads to a downstream cascade signaling activity. DDRs thus play an important role in modulating collagen synthesis and organization during scar formation. As a corollary, we hypothesized that differential expression of collagen and DDRs may determine the nature and extent of tissue scarring.
As a part of a fundamental assessment of receptor activity, the expression levels of DDR1 between normal human dermal fibroblasts and keloid fibroblasts were compared using immunohistology, Western blot, and Northern blot. Next, blocking DDR1 expression in keloid fibroblasts by transfection of DDR1 antisense oligonucleotides to compare the expression of downstream signal molecules in mRNA expression levels, the inhibition of proliferation of keloid fibroblasts, and collagen production through [3H] proline incorporation assay, was evaluated.
Materials and Methods
Tissue culture
Full-thickness skin specimens of keloids were obtained by surgical excision in 10 patients (age range 20–65 years). All patients gave written informed consent, as approved by the Ruijin Hospital Ethics Committee. The samples were removed from the chest (2 patients), earlobe (3 patients), back (4 patients), shoulder (2 patients), and lower extremities (1 patient). No prior keloid treatment had been performed. Fibroblast cultures analyzed in this study were from explanted specimens. Normal human dermal fibroblasts were obtained from 3 patients (age range 21–65 years) who had undergone cosmetic surgery. The samples were obtained from the chest in two patients and the face in the third. All patients were healthy and had no other illnesses. All fibroblast cell lines were derived from primary tissue explants and maintained in 10% fetal bovine serum and Dulbecco’s modified eagle medium ([DMEM], HyClone Media, Logan, UT) at 37˚C in a humidified incubator with 5% CO2. Upon confluence, cells were washed twice in phosphate-buffered saline and split 1:3 using 0.25% trypsin. Only cells from less than the 7 passages were analyzed in this study.
Immunohistology
Immunohistology of skin specimens harvested from different patients was performed according to the protocol. The skin specimens were fixed in 10% buffered formalin for 24 hours at room temperature then processed and embedded in paraffin. The specimens were then de-waxed in xylene for 30 minutes before immersion in 3% hydrogen peroxide solution for 20 minutes at room temperature. The solution was placed in citrate and heated at 37˚C for 10 minutes for antigen retrieval. The solution then was cooled in buffer at room temperature for 20 minutes and blocked with 5% BSA for 20 minutes at room temperature. Diluted DDR1 antibody ([1:50, sc-532] Santa Cruz Biotechnology, Inc., Santa Cruz, CA) was added to the solution, and allowed to set for 3 hours at room temperature. The solution was covered with biotinylated goat anti-polyvalent secondary antibody and set at room temperature for 30 minutes. The solution then was covered with drops of streptavidin-horseradish peroxidase (HRP) and mixed with 0.05% DAB and 0.03% hydrogen peroxide. This solution was placed in a hematoxylin bath and washed for 2 minutes. Control immunohistology was performed using non-specific goat IgG.
RNA extraction and cDNA synthesis
Total RNA from fibroblasts was extracted using TRIzol according to the manufacturer’s protocol (Gibco-BRL Life Technologies, Invitrogen, Gaithersburg, MD) before reverse transcriptase (RT)-PCR was performed.9 Complementary DNA was synthesized from 5 μg total RNA using avian myeloblastosis virus (AMV) reverse transcriptase (Promega Corporation, Madison, WI) and random primers.
Conventional polymerase chain reaction
For PCR amplication, 2-μL of cDNA with 0.5-μL Ex Taq DNA polymerase (Takara, Japan), 4-μL dNTP, and 1-μL of each primer were used in a final volume of 50 μL. The PCR primers for DDR1, Col-1A, Col-3A, and glyseraldehyde-3-phosphate dehydrogenase (GAPDH) were designed using OLIGO primer analysis software version 5.0 (NBI, Plymouth, UK). The DDR1 upstream primer was GGACATACCGTGGGCGGACT, and downstream primer was CCATGCGCCACAACCTAGG (Genbank code NM_001954.4). The Col-1A upstream primer was GGCGGCCAGGGCTCCGACCC, and downstream primer was AATTCCTGGTCTGGGGCACC (Genbank code NM_000088.3). The Col-3A upstream primer was TGGTGTTGGAGCCGCTGCCA, and downstream primer was CTCAGCACTAGAATCTGTCC (Genbank code NG_007404.1). The GAPDH upstream primer was TCATTGACCTCAACTACATG, and downstream primer was AGTGATGGCATGGACTGT (Genbank code NM_002046.3). The PCR products were visualized with agarose gel electrophoresis. Densitometric values for all mRNA samples were normalized according to values for expression of GAPDH. All experiments were performed twice.
Western blot analysis
Immunoblotting was performed as previously described.10 Briefly, keloid and normal human dermal fibroblast cultures containing the same number of cells (1 x 106) were rinsed twice with cold phosphate-buffered saline and then lysed with a cold-lysed buffer—radio-immunoprecipitation assay (RIPA) buffer. To this solution, 150 mM NaCl, 1% NP-40, 0.1% SDS (sodium dodecyl sulfate), 50 mM Tris-HCl, pH 7.4, 0.25% sodium deoxycholate, and 1 mM phenylmethylsulfonyl fluoride (PMSF) were added and allowed to set for 1 hour. Cell lysates were then scraped from dishes, repeatedly pipetted to shear DNA, and allowed to incubate on ice for 30 minutes before centrifugation at 10,000 rpm for 10 minutes to remove insoluble material. Protein concentrations were determined by Bio-Rad assay (Bio-Rad Laboratories, Inc., Hercules, CA). Equal amounts (30-μg) of protein samples were boiled for 5 minutes and then electrophoresed in 10% sodium dodecylsulfate- polyacrylamide gels (SDS-PAGE). These samples were transferred to cellulose acetate membrane (CAM) by using a semi-dry electro-blotting device (Yrdimes, Wealtec Corporation, Sparks, NV). Membranes then were blocked with Tris-buffered saline (TBS) containing 5% fat-free milk for 12 hours. Subsequently, the membranes were rinsed in 0.1% polysorbate-20 (Tween) three times for 10 minutes each, and then incubated with the appropriate primary antibody at 4˚C overnight. The primary antibodies included DDR1 (Santa Cruz Biotechnology, Inc., Santa Cruz, CA) and β-actin. After the blots were washed in 0.1% polysorbate-20 three times, they were incubated with an IRDye® 800 conjugated affinity purified IgG ([H&L], Rockland Immunochemicals, Gilbertsville, PA) for 50 minutes. Next, the membranes were washed with TBS containing 0.1% polysorbate-20 three times. The Odyssey Infrared Imaging System was used for signal detection (LI-COR, Lincoln, NE).
Antisense and nonspecific oligonucleotides transfection
The sequence of the DDR1 antisense oligodeoxynucleotide (ASODN) was 5'-GCCTCTGGTCCCATAGC-3' complementary to bases 331-347 of DDR1. The sequences 5'-TCTGGTGCCTACCTATG-3' were used as nonspecific oligonucleotides (NSODN). The oligonucleotides used were end-phosphorothioated and synthesized at Shanghai Sangon Biotechnology Company, Ltd. (Shanghai, China). To confirm the above gene sequences as unique (not consistent with other gene sequences), a search of the Basic Local Alignment Search Tool (BLAST) IMAGE EST (Expressed Sequence Tags) library was conducted.
These oligonucleotides were transfected into keloid fibroblasts cultured with lipofectamine reagent (Gibco-BRL Life Technologies, Invitrogen, Gaithersburg, MD) according to the manufacturer's instructions. The final concentration of oligonucleotides was adjusted to 1 mmol/L. The final concentration of lipofectamine was 10 mmol/L. The experimental groups were divided into low, medium, and high concentration of oligonucleotides. In the low-concentration group, DDR1-ASODN was added into 9.6 μg. In the medium-concentration group, 19.2 μg was added. In the high-concentration group, 38.4 μg was added. A defined number of cells (3×105 to 5×105) were seeded in 25-cm2 flasks. These cells were 80% to 90% confluent at the time of transfection. Antisense or nonspecific oligonucleotides and lipofectamine reagent were mixed and preincubated for 15 minutes at room temperature then mingled for 20 minutes. The final volume of oligonucleotides mixture in each vessel was 10 mL. Keloid fibroblasts were treated with the oligodeoxynucleotide mixture at 37˚C in a CO2 incubator until ready to assay.
Collagen synthesis
Keloid fibroblasts were seeded in 24-well microculture plates at a density of 1 x 105 cells/well with 1-mL DMEM (HyClone Media, Logan, UT) supplemented with 10% fetal bovine serum. By day 3, the cells were confluent and the transfection procedure was conducted according to the manual and design. Twenty-four hours after transfection, media were removed and replaced with 1-mL DMEM supplemented with 0.5% fetal bovine serum and L-ascorbic acid (Sigma-Aldrich, St. Louis, MO). Twenty-four hours later, [3H] proline (10 mCi/mL) was added to 0.5 M acetic acid containing pepsin. Following incubation overnight at 4˚C, extracellular [3H]-labeled collagen was extracted and purified by successive salt precipitation at acid and neutral PH. This was followed by precipitation in 20% ethanol. Final precipitate was solubilized in 0.5-M acetic acid and placed in scintillation liquid. Radioactivity levels were determined. Results were averaged from three identically treated wells and reported as counts per minute (cpm).
Analysis of cell proliferation inhibition
The number of viable cells was determined by the 0.4% Trypan blue dye (Sigma-Aldrich, St. Louis, MO) exclusion test. Percentage of inhibition of cell proliferation was calculated by the following formula: Inhibition % = [(N-NT)/N] 100%, where N is the number of untreated cells and NT is the number of treated cells.
Quantification of blots
Densitometric analysis was performed using image analyzer software, the FluorChem™ FC 800 system (Alpha Innotech, San Leandro, CA), which measures the sum of all pixel values after background correction.
Statistical analysis
All data are expressed as mean ± SE of at least three independent experiments. One-way analysis of variance was used to test for statistical difference. Statistical significance was set at P < 0.05.
Results
Immunohistochemistry in keloid skin
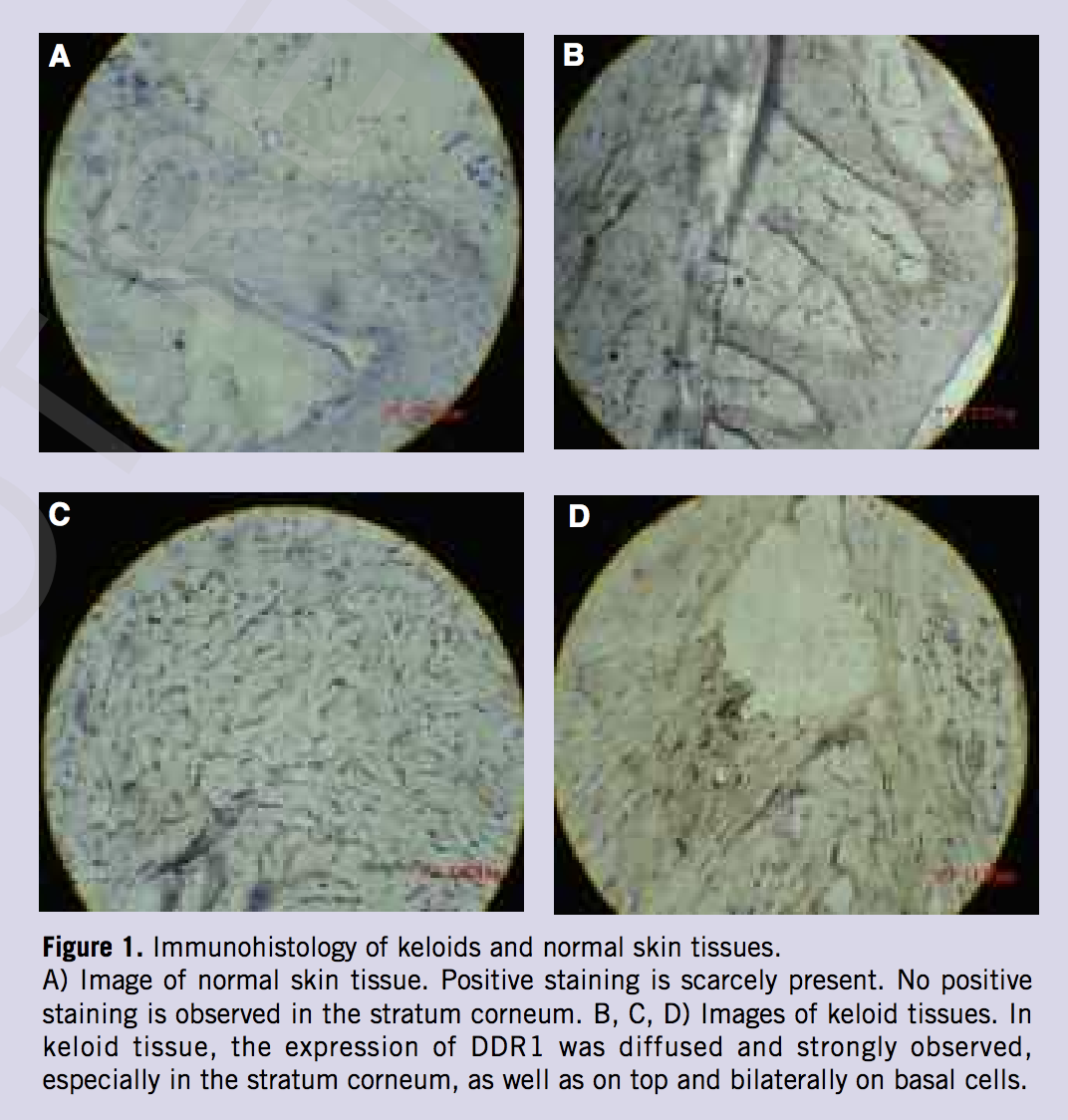
In keloid skin sections, vast and strong overexpression of DDR1 was observed, especially in the stratum corneum including the top and bilateral sides of basal cells. Positive staining that was aggregated and clumped in the cytoplasmic margin of the cells was observed. However, in normal skin tissues positive staining was scarcely present (Figure 1).
Western blot analysis
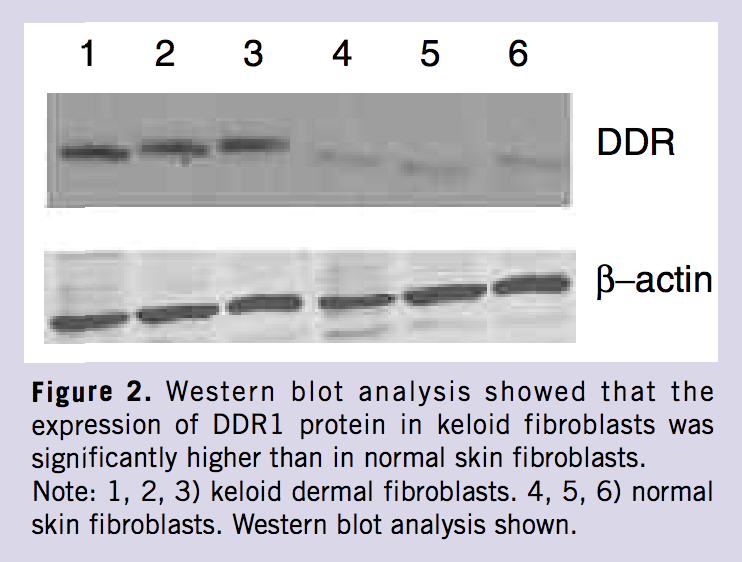
Keloid fibroblasts from three patients and normal skin fibroblasts from three healthy people were examined by Western blot. The results showed that the expression of DDR1 was highly increased compared with the normal skin fibroblasts (Figure 2).
Inhibition of cell proliferation
Keloid dermal fibroblast viability was decreased significantly by DDR1-ASODN treatment in a dose-dependent manner. Additionally, keloid dermal fibroblast death was increased.
Expression of DDR1, Col-1A, and Col-3A after transfection
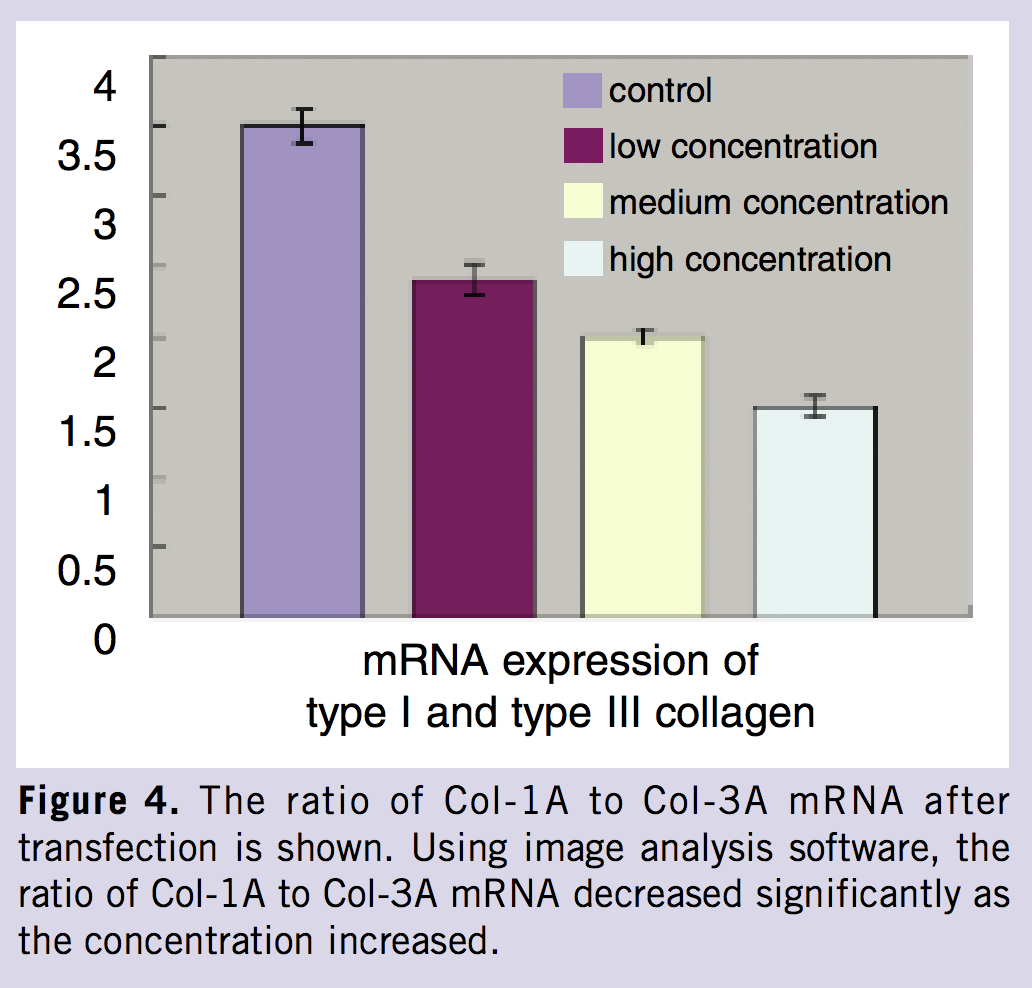
Examining the expression of Col-1A and Col-3A was sought initially, due to the close relationship between DDR1 and collagen. After transfection, Western blot analysis showed that the relative expression of DDR1 protein was inhibited significantly after lipofectamine-mediated DDR1-ASODN transfer into fibroblasts. The expression of Col-1A and Col-3A also was inhibited. The extent of inhibition was positively related with transfection concentration. There were no significant differences between control groups. Reverse transcriptase-polymerase chain reaction also showed that the expression of DDR1 was inhibited significantly after transfection. The relative expression of α (I) collagen and α(III)-collagen mRNA was inhibited significantly in the experimental groups compared with control cells (Figures 3, 4). Note the progressive decrease in mRNA levels as the concentration increased. The ratio of α (I) to α (III)-collagen mRNA decreased significantly as concentration increased.
Total collagen synthesis by keloid fibroblasts after transfection
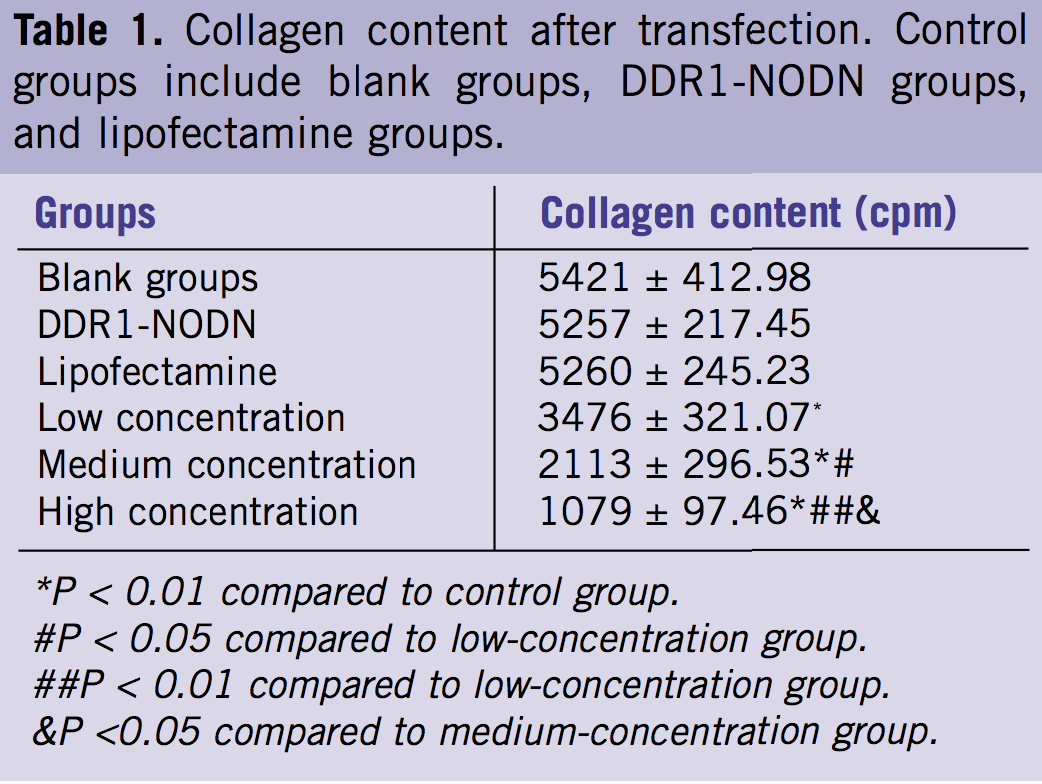
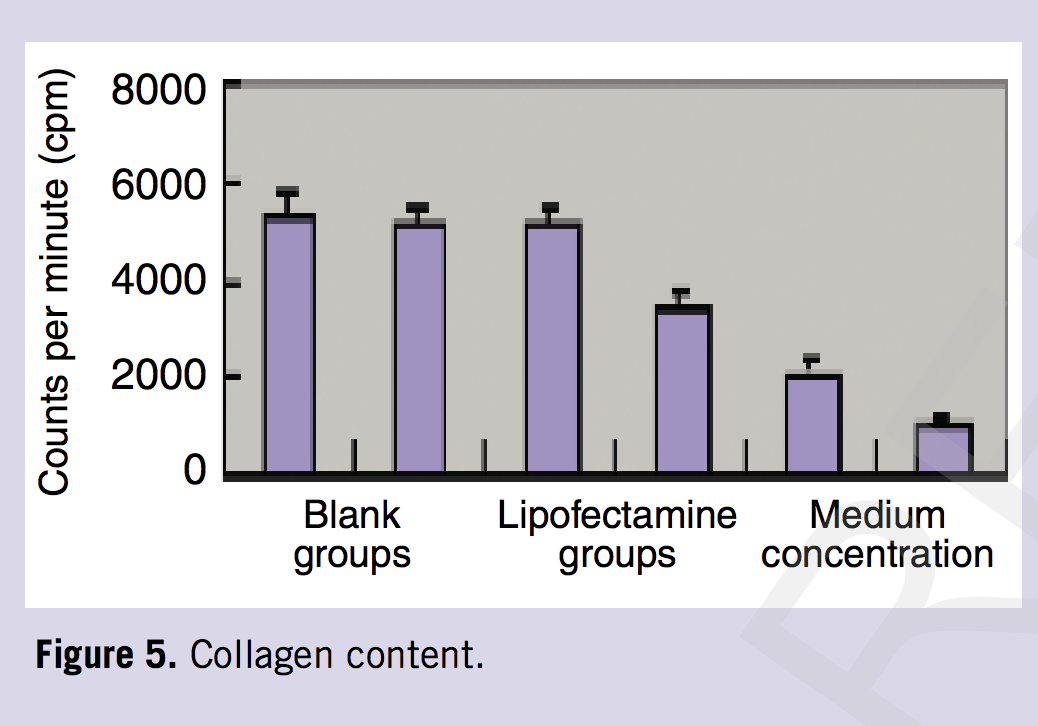
To determine the level of collagen protein produced by keloid fibroblast after transfection, total collagen production by [3H] proline incorporation was examined. The findings revealed lower collagen production in the experimental groups after transfection, and higher collagen production in the control groups. The extent of decrease became more significant as the concentration increased (Table 1, Figure 5; P < 0 .05, P < 0.01).
Discussion
Keloids are benign dermal tumors that form during an abnormal wound-healing process in genetically susceptible individuals.11 They are clinically defined as scars that extend beyond the original wound margin and are characterized by excessive deposition of collagen in the dermis as a result of dysregulated response to cutaneous wounding. They pose a difficult problem to plastic surgeons because most treatments are largely ineffective. Appropriate wound healing involves the complex regulation of many factors, including various cells, cytokines, growth factors, and ECM in a complex milieu of both local and systemic factors. The structure and configuration of collagen are believed to play critical roles in successful wound healing. Hence, regulation of collagen is fundamental in tissue remodeling and represents a crucial step in the process of wound healing with or without scar formation.
The recently discovered discoidin domain receptor (DDR1 and DDR2) group is one of three collagen receptor groups that modulate production and organization of collagen in scar tissue. Functionally, DDRs are tyrosine kinase receptors, which are distinct from other receptor groups due to their extracellular domain that resembles discoidin8—a developmentally regulated lectin first described in the slime mold dictyostelium discoideum.12,13 Many studies have revealed that DDR1 is overexpressed in human tumors including breast, ovarian, esophageal, and brain tumors and are believed to be a tumor marker14; the reason is the expression of DDR1 were highly up-regulated as collagen receptor, which sense the actively remodeling of extracellular matrix, especially the collagen, resulting from the powerful erosion of malignant tumors. The authors hypothesize that DDR1 may be highly expressed in keloids for its excessive deposition of collagen and dermal tissue remodeling.
In this study, the role of DDR1 in keloids was evaluated. As expected, histological analysis demonstrated that the expression of DDR1 was widely and strongly distributed in keloid tissues. The extent of expression was highly increased on the keloid margins and less increased in the center. This showed that the expression of DDR1 was related to keloid infiltration. The authors assume that in the area of infiltration the higher extent of destruction in the ECM was related to the increased ECM remodeling and higher DDR1 expression. The expression of DDR1 was also higher in the normal skin around the keloid than the normal, healthy skin (data not shown), revealing that the person was genetically susceptible due to the keloid. However, in order to avoid new keloid formation, we cannot draw normal skin samples from patients already affected by keloid.
Additionally, when the cell culture was made from the tissue explants, the keloid fibroblasts were more easily cultured from the keloid margin. Thus, the keloid fibroblasts were from the area of keloid infiltration. In order to create further experiments, the expression of DDR1 was made by Western blot analysis between keloid fibroblasts and normal skin fibroblasts. The result showed that the expression of DDR1 was significantly higher in keloid dermal fibroblasts than in normal skin fibroblasts, demonstrating a difference between the function and phenotype of keloid fibroblasts and the normal skin fibroblasts. Others have also demonstrated this phenomenon.11
Next, we examined the production of collagen in keloid dermal fibroblasts after blocking the expression of DDR1. After liposome-encapsulation with DDR1-ASODN transfected into the keloids dermal fibroblasts, the relative expression of DDR1 mRNA was dramatically inhibited. This inhibition progressively increased with an increased concentration of DDR1-ASODN. The relative expressions of Col-1A and Col-3A mRNA levels also were significantly inhibited. Inhibition gradually increased as the final concentration of DDR1-ASODN increased. Using image analysis software, it was observed that the ratio of α (I) to α (III)-collagen mRNA decreased significantly as the concentration increased. Moreover, the production of collagen also decreased significantly after transfection. The extent of decrease continued as the concentration of DDR1-ASODN increased. These results suggest that blocking the expression of DDR1 would downregulate behavior of keloid dermal fibroblasts. However, this conclusion does not completely support the results of Chin et al7 who found that there was a gradual increase in collagen production that was concomitant with progressively lower expression of DDR1 in fetal rat fibroblasts during the characteristic fetal rat skin conversion from scarless phenotype to a scarring phenotype. The reason why this occurs may be because during morphogenesis the tissue and organs build up and the characteristic fetal skin has scarless phenotype in which the DDR1, as sensors for ECM, is highly expressed to facilitate collagen modulation and organization. As the fetal scar healing period progresses, the miniature of an adult form resolves and the refined modulation of collagen organization gradually reaches homeostasis, and as a result, the expression of DDR1 is gradually decreased. Once the skin is wounded, this tissue building machinery is dramatically reawakened and the repair becomes a recapitulation of morphogenesis. As a result, ECM forms and the expression of DDR1 is greatly increased.15 However, this process is different in keloid formation, hence the variation in findings.
Additionally, it was observed that the viability of keloid dermal fibroblasts significantly decreased after treatment with DDR1-ASODN. Ongusaha et al16 also reported that DDR1 activated the mitogen-activated protein kinase (MAPK) cascade. Additionally, Ongusaha et al suggest that DDR1 can function as a survival effect directly. Their data support our finding that blocking the expression of DDR1 is associated with decreased cell viability.
Furthermore, according to the manual of Lipofectamine™ 2000 transfection reagent (Gibco-BRL Life Technologies, Invitrogen, Gaithersburg, MD), transfection efficiency is only 48% in primary human fibroblasts. The manual suggests that only half of keloid dermal fibroblasts can successively interrupt DDR1 expression in the culture plate. The results of the present study further demonstrate that some DDR1 can exert regulatory effect on collagen structure. These findings reveal that DDR1 can sense the quality of collagen, activate the signals, and regulate cellular behavior thus modulating collagen proliferation, production, and organization.
The distribution of DDR2 is not as widely dispersed as that of DDR1. In our experiments, the expression of DDR2 remained fairly constant between keloid fibroblasts and normal dermal fibroblasts (data not shown). The exact role of DDR2 will require further investigation.
Another issue that needs to be studied further is how these in-vitro findings correlate with in-vivo results. Additionally, it needs to be addressed why a difference exists in healing at two different sites in a person who suffers from keloid formation at a given anatomical location. A follow-up study using different skin samples—normal and keloid tissue from the same subjects—will shed some light on the differences in fibroblast heterogeneity. We believe that this study will provide baseline data that can be used in future clinical trials and also shed some light on other unresolved issues.
Conclusions
This study has shown for the first time that keloids can be inhibited by targeting DDR1 signaling using a DDR1-ASODN treatment. The effect of DDR1-ASODN treatment on long-term results and animal model remains to be investigated. However, this study demonstrates a close relationship between collagen and DDRs in keloid dermal fibroblasts and is likely to open newer avenues for further investigations on wound healing, which may prove to be useful in the future.
Acknowledgments
The authors would like to thank Dr. C-Y Ni and Y-H Cao for their technical assistance on this project.
Address correspondence to: Yuzhi Jiang, MD, PhD and Xin Xing, MD, PhD, Shanghai Burn Institute, Ruijin Hospital Shanghai Jiaotong University, 197 Ruijin No.2 Road, Shanghai 200025, P. R. China; Email: yuzhijiang@hotmail.com
References
Sign Up Today





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
