

Changes of Transforming Growth Factor-β1 and Extracellular Matrix in the Wound Healing Process of Rats Infected With Pseudomonas aeruginosa
Abstract
Objective. This work aims to investigate the changes of transforming growth factor-β1 (TGF-β1) and extracellular matrix (ECM) proteins in rats with chronic skin ulcers infected with Pseudomonas aeruginosa. Methods. Forty-eight female Wistar rats, aged 8 weeks, were randomly divided into a simple cutaneous wound group (group A) and a cutaneous wound plus Pseudomonas aeruginosa inoculation group (group B). On postoperative days 1, 3, 7, and 10, hematoxylin and eosin staining, real-time polymerase chain reaction, immunohistochemistry (IHC), and enzyme-linked immunosorbent assay (ELISA) methods were used to detect the epithelization rate, the mRNA, and the protein expressions of matrix metalloproteinases (MMP) 2, MMP-9, tissue inhibitors of metalloproteinases (TIMP)-1, and TGF-β1. Results. Compared with group A, the expression of TGF-β1 and the epithelization rate were delayed in group B. At 3 days postoperation, the expression of collagen III was reduced in group B. At 7 days postoperation, MMP-9 had an extremely high expression but collagen and TIMP-1 had a low expression. Seven days after being infected with Pseudomonas aeruginosa, the expression of TGF-β1 was reduced in the wounded skin of rats, and the dissolved ECM was much more than that synthesized in wounds. Conclusion. The authors’ experiments suggest the wounded skin tissue infected with Pseudomonas aeruginosa delayed the expression of TGF-β1, and then caused a change in the ECM biology relevant to fibrosis and regeneration, thus affecting the wound healing process.
Introduction
Chronic wounds, defined as those that do not heal in an orderly set of stages and in a predictable amount of time,1 are often accompanied by infection, especially in wounds such as skin ulcers, pressure ulcers, burns, and diabetic foot ulcers. Pseudomonas aeruginosa is one the most common clinical pathogens and is also a kind of lethal opportunistic infection.2 Chronic infection usually occurs when the patient’s resistance is low, which not only increases the burden on patients but also severely reduces the speed and quality of the healing process.3 The infecting bacteria could competitively absorb nutrition from wounds and secrete toxins to destroy the wound surface, thus delaying healing. The bacteria also causes abnormal cytokine secretion from host cells. The changed expression of growth factors also influences the healing of wounds to different degrees. To date, growth factors are widely applied on the treatment of refractory cutaneous wounds. Transforming growth factor-β1 (TGF-β1) is a widespread cytokine and has a variety of biological activities. It plays an important role in regulating the generation of fibrous tissue and the synthesis of extracellular matrix (ECM). It is a growth factor closely related to the healing of wounds and affects almost all phases in the wound healing process.4 Matrix metalloproteinases (MMPs) are an important family involved in the healing process whose members could modulate inflammation, degrade the ECM, and promote cell migration and remodeling. However, excessive MMPs will lead to the development of chronic wounds. In this study, the authors used Pseudomonas aeruginosa-infected wounds to investigate the relationship between the formation of chronic refractory cutaneous wounds caused by Pseudomonas aeruginosa and the expression of TGFβ1, MMP-2, MMP-9, and its inhibitor TIMP-1, to provide a theoretical basis for the clinical treatment of refractory wounds
Materials and Methods
Animals. Forty-eight female Wistar rats, aged 8 weeks of age and weighing 220 g to 310 g, were housed in a limited-access rodent facility with a temperature of 24°C ± 2°C, a humidity of 30%-70%, and a 12-hour light/dark cycle. Only healthy rats were used for the experiment. This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The animal use protocol has been reviewed and approved by the Committee on the Ethics of Animal Experiments of the Jilin University.
Preparation of Pseudomonas aeruginosa. The Pseudomonas aeruginosa strain used in this study was wild-type PAO1, obtained from the Department of Microbiology of Jilin University, Changchun, China. Fresh bacterial suspension was inoculated onto a sheep serum plate (Agrisera, Vännäs, Sweden) and incubated at 37°C for 24 hours. Colonies in good growth states were trans-inoculated into brain-heart infusion medium and incubated at 37°C for 24 hours to an optical density at 600 nm of 0.6 to 0.8, which corresponds to 108 colony-forming units (CFU)/ml at the mid-log phase. The cultured cells were centrifuged 2 times in phosphate buffered saline (PBS) at 2,000 rpm and then suspended in PBS at a concentration of approximately 3.6 × 109 CFU/ml. Bacterial suspension (14 µl) were then inoculated into the base of the wounds (n = 48) at a concentration of 5 × 107 CFU/wound. Two wounds were created on each rat, with 48 wounds used for each group (96 wounds in total); wounds without inoculation of bacteria served as the blank control.
Animal grouping and model building. Rats were randomly divided into 2 groups: Simple cutaneous wound group (group A) and cutaneous wound plus Pseudomonas aeruginosa inoculation group (group B). All rats were anesthetized with 10% chloral hydrate by intraperitoneal injection and the backs of the animals were shaved to expose the skin after being disinfected by 10% iodophors. Under the aseptic condition, the back skin was cut by a 6 mm diameter biopsy punch and full-thickness wounds were created using sterile surgical scissors. Two wounds were created on each rat. In group B, bacterial suspension (14 µl) was inoculated into the base of the ulcer at a concentration of 5 × 107 CFU/wound. The periphery of each wound was sealed with a hydrocolloid dressing (DuoDerm Extra Thin dressing, ConvaTec, Skillman, NJ). The entire area of the wound was then covered with polyurethane film to ensure an enclosed environment. The day on which the wound was made was considered day 0. Acetamidophen 10 mg/kg was given for pain control during the entire procedure.
Determining the bacterial count and infection. On days 1, 3, 7, and 10 after infection, photographs of the rats were taken to record appearance and signs of inspection in the cutaneous wounds. Exudate quantity was estimated and collected by a sterile cotton swab and diluted tenfold, then sowed into sheep serum medium for 24 hours. Bacteria colonies were then counted using the previously described method. The entire layer of wounded skin was cut out with a sterile scalpel blade and put into a sealed tissue grinder containing 10 ml saline. After the tissue was completely ground, it was diluted tenfold and cultured onto the sheep serum plate for 24 hours at 37°C. Afterwards, the colonies were counted and recorded.
Detection of epithelization. Three pieces of wounded skin were gathered from both groups on days 1, 3, 7, and 10 after the inoculation with the bacteria. Each specimen was vertically divided into 2 pieces with a blade. One piece was fixed with 10% formalin and embedded in paraffin, and the other was used for quantitative polymerase chain reaction (Q-PCR). The skin of the wound was cut in 5 µm and stained with hematoxylin and eosin (HE) and then observed under light microscopy to determine the epithelialization level and contraction rate. This could be formulated as follows:
Epithelialization level = [total wound surface area (mm2) - residual wound area (mm2)] / total wound surface area (mm2)
Wound contraction rate = [initial wound area (mm2) - post-injury area (mm2)] / initial wound area (mm2)
Detection of TGF-β1 expression using enzyme-linked immunosorbent assay. Wounded tissue cut from both groups was weighed and cut into pieces using scissors. The tissue was then diluted tenfold and ground using a tissue grinder. Afterwards, the homogenate was centrifuged at 3000 rpm for 3 minutes and the supernatant was collected for enzyme-linked immunosorbent assay (ELISA). Transforming growth factor-β1 was detected by conventional sandwich ELISA using TGF-β1 ELISA kit (Sigma-Aldrich, St. Louis, MO), according to the manufacturer’s instruction. The optical density was measured at 450 nm on a spectrophotometer.5
Quantitative polymerase chain reaction. Quantitative real-time polymerase chain reaction (TaqMan, Invitrogen, Life Technologies, Grand Island, NY) was performed using a real-time Q-PCR kit (SYBR Premix Ex Taq, Takara Biotechnology [Dalian] Co, Dalian, China). Total RNA was reverse transcribed to cDNA using Superscript II reverse transcriptase (Invitrogen, Life Technologies, Grand Island, NY) and oligonucleotide primers. Quantitative real-time PCR (RT-PCR) analysis of gene expression was performed in a 25µl reaction volume containing cDNA, SYBR Premix Ex Taq (Takara Bio Inc, Shiga, Japan), TaqMan Universal PCR Master Mixture and primers for each gene. Gene-specific primers for rat MMP-2, MMP-9, and TIMP-1 were obtained from Sangon Biotech Co, Ltd (Shanghai, China) (Table 1). The relative quantitative method was used for the quantitative analysis and fold change (FC) was used to present data. The reaction conditions were as follows: 95°C denaturation for 10 minutes, 1 cycle; then 95°C denaturation for 30 seconds, 58°C annealing 40 seconds, and 72 °C extension 40 seconds for 45 cycles. Fluorescence signals were measured after 45 PCR cycles, and all samples were normalized to β-actin housekeeping gene levels. All samples were run in duplicate. The relative quantitative method was used for the quantitative analysis and FC was used to present data.
Immunohistochemistry analysis. Serial sections of paraffin-embedded specimens were performed and then stained by HE immunohistochemistry method (Santa Cruz Biotechnology, Inc, Dallas, TX) in the standard protocol. Phosphate buffered saline was used as a negative control. Positive results were determined according to the national immunohistochemical techniques and diagnostic criteria.6
Cytoplasmic positive or nuclear positive cells were determined in brownish yellow or dark brown yellow. Five high-power fields (400x) of each slice were observed. A rate > 25% was determined as positive samples and < 25% or zero positive cells was regarded as negative. The positive cells also could be divided into weak, moderate, and strong according to their dye degrees. Positive cells that were yellow, or with a positive cell rate ranging from 25% to 50%, were regarded as weakly positive. Positive cells that were brownish yellow, or with a positive cell rate ranging from 50% to 75%, were regarded as moderately positive. Positive cells that were dark brown-yellow, or with a positive cell rate > 75%, were considered strongly positive.
Statistical Analysis. Statistical analysis was performed with the SPSS software program v 13.0 (SPSS Inc, Chicago, IL). All results were presented as means ± standard deviation. Differences between the 2 groups were compared using 2 sample t tests. For all tests, P < 0.05 was considered significant.
Results
Identification of animal model. The 2 groups of wounds were examined by visual observation, and no obvious redness or swelling were found in the simple cutaneous wound group (group A). In the bacterial inoculation group there was more yellow-green and odorous exudate and significant swelling around the wound on postoperative day 2. Bacterial number in each wound was more than 106 CFU, meeting the standard for infection. The number of bacteria in the infected tissue was more that 105 CFU. The HE staining results showed that in the control group, neutrophils increased gradually and peaked at day 3, then gradually decreased; however, in the infected group, neutrophils increased sharply on the first postoperative day and then gradually reduced with time.
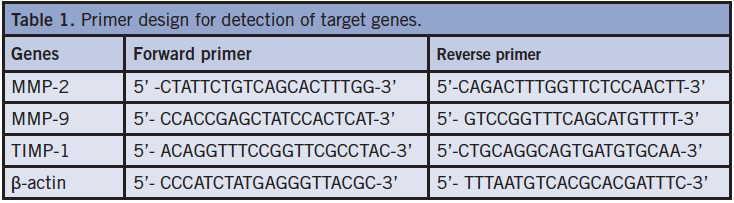
Epithelization level of wounds in both groups. Wounded skin in group A appeared epithelialized in the first 3 days when observed under an optical microscope. The epithelialization level and wound contraction rate reached 83% and 27.3%, respectively, on postoperative day 7, and then 95.6% and 63%, respectively, on postoperative day 10, which was close to complete epithelialization. In group B, the epithelialization level was only 50% and the rate of wound contraction was 30.3% at postoperative day 7. On postoperative day 10, the epithelialization level was almost close to group A and the wound contraction rate was 45% (Figure 1).
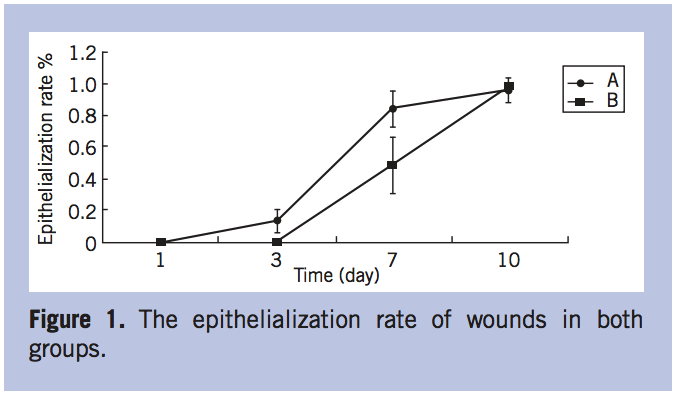
The TGF-β1 expression in wounded skin. The expression of TGF-β1 in the wounded skin had an upward trend postoperatively in both groups. It appeared significantly higher in group B than in group A at 7 days post-infection (P < 0.05). In group B, the TGF-β1 expression at 7 days post-infection was much higher on day 1 (P < 0.05) (Figure 2). The expression of TGF-β1 was not significantly different between the 2 groups at 10 days post-infection (Figure 2).
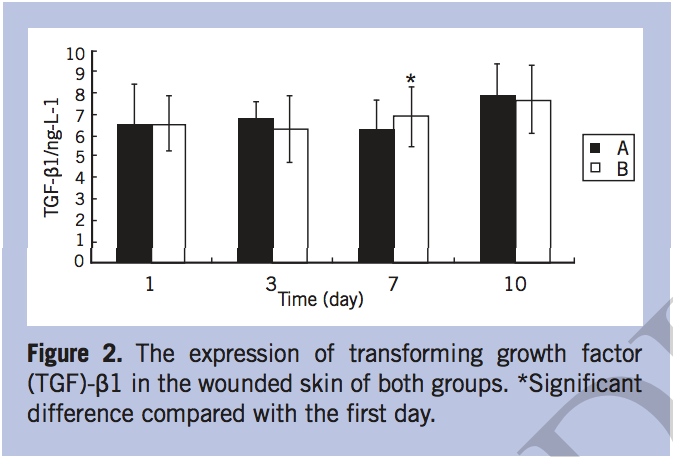
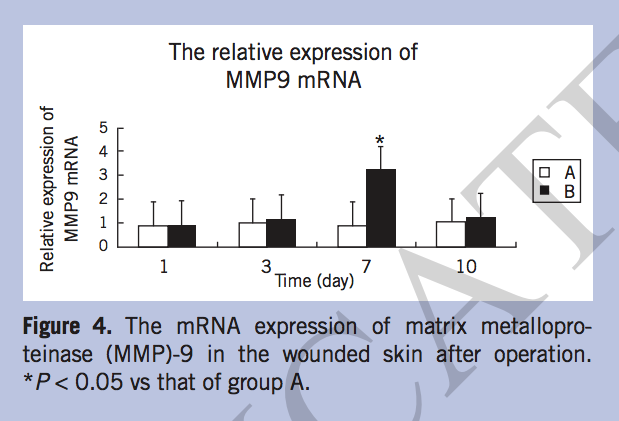
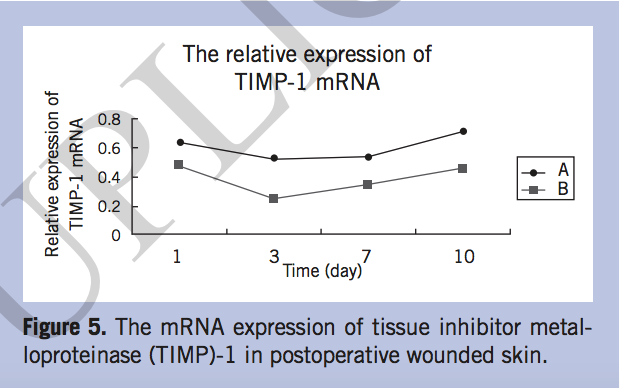
The mRNA expression of MMP-2, MMP-9, and TIMP-1. The mRNA expression of MMP-2 increased gradually after operation in both groups, but increased more slowly in group B than in group A. On postoperative days 3, 7, and 10, the MMP-2 mRNA of group A elevated significantly (P < 0.05) (Figure 3). The expression of MMP-9 in group A was lower than that of group B, especially on day 7, when MMP-9 mRNA increased sharply in group B (P < 0.05) (Figure 4). The relative expression of TIMP-1 mRNA in group A was significantly higher than that in group B on day 7 (P < 0.05) (Figure 5).
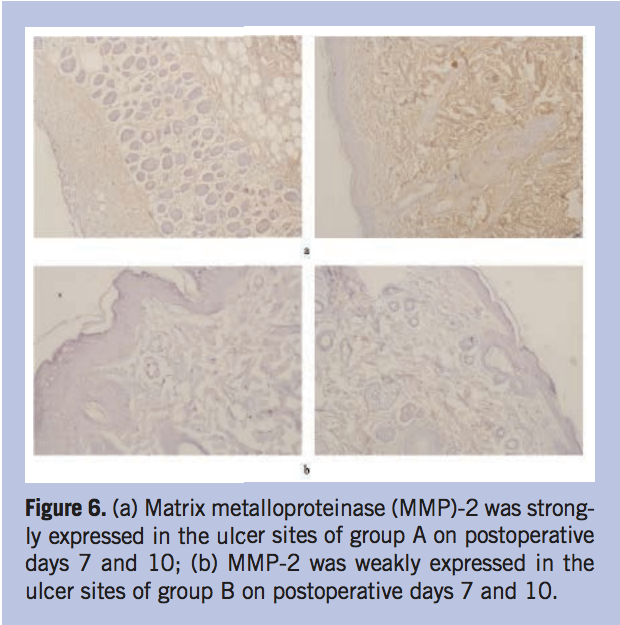
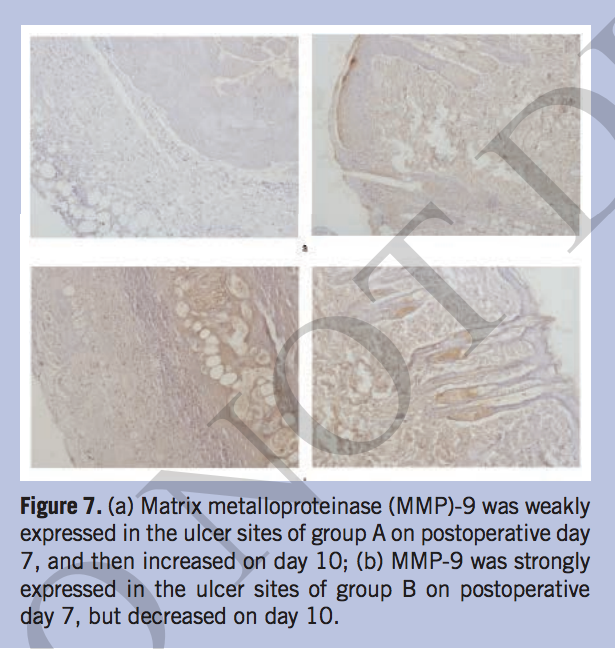
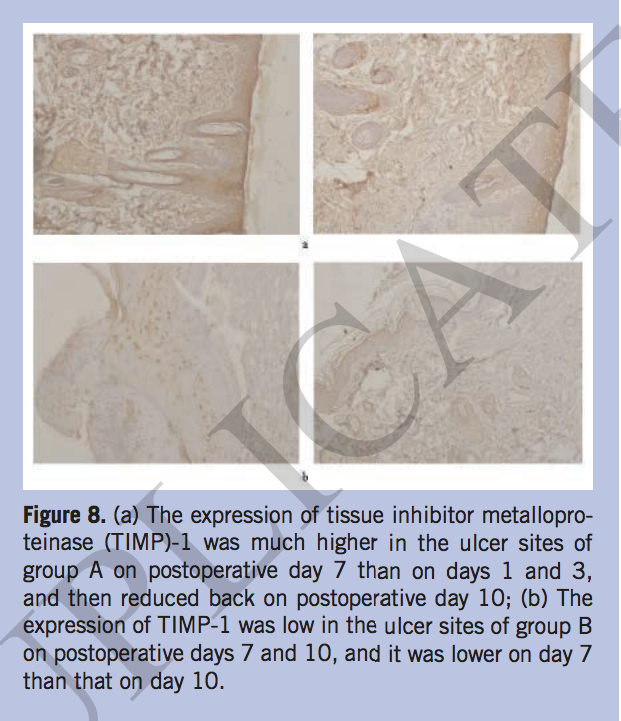
The expression of MMP-2, MMP-9, and TIMP-1 in situ. The authors also detected the expressions of MMP-2, MMP-9, and TIMP-1 using immunohistochemistry (IHC) method to further investigate the wound repair situation. The IHC results showed that MMP-2 was strongly expressed in the ulcer site of group A on postoperative days 7 and 10. However, it was only occasionally positive in group B during the same time period (Figure 6). The expression of MMP-9 in group A was weak at postoperative day 7 and then increased at day 10. In group B, MMP-9 was expressed at a higher level on day 7 compared to days 1 and 3, and then decreased at day 10 (Figure 7). Tissue inhibitor metalloproteinase-1 had the strongest expression at 7 days postoperation, decreased at 10 days in group A, and TIMP-1 had a lower expression in group B than group A on postoperative day 10. (Figure 8).
Discussion
Wound healing is a complex process dependent on epithelialization, the formation of granulation tissue, the modeling of the ECM, and scar formation.7,8 To date, there is no single molecular signal that could be used to indicate the wound healing process is progressing.There are a large number of factors that can delay wound healing, especially when microorganisms are present in a wound such as continuous production of inflammatory mediators, metabolic waste, oxygen free radicals, toxic substances, the sustaining of neutrophil activation status, and the production of cytolytic enzymes. These factors prolong the inflammatory response and the process of wound healing.2 In addition, the bacteria present in the wound compete with the healthy tissue for available nutrients, resulting in tissue hypoxia, which could cause granulation tissue weakening and bleeding, thus reducing the number of fibroblasts and the production of collagen, consequently damaging the epithelialization process. Bacterial infection can also produce subclinical tissue damage. Studies9,10 have shown that if the bacterial number at the wound exceeds 105 or 106/g, it can cause local lesions or delay wound healing. After becoming infected with Pseudomonas aeruginosa, more exudates are produced in the wound, leading to the increased skin tension and dehiscence of the regional wound. Meanwhile, the combined effect of endotoxin, exotoxin, and the proteolytic enzyme also causes the increasingly biological effects of the cytokine and free radical damage, resulting in tissue edema, hemorrhage, increased purulent discharge, and proteolysis. The growth of granulation tissue slows down the repair of injury cavity and seriously affects the speed of wound healing.7,11 It has been reported that the dissolution of corneal stroma is due to the excessive degradation of corneal ECM proteins, collagen type I, and proteoglycan. Infiltration of white blood cells is involved in this process. Polymorphonuclear leukocytes can cause pathological tissue damage by releasing free radicals, elastase, cathepsin, collagenase, and gelatin enzymes. The multicore elastase can degrade many ECM, elastic protein, collagen, and fibronectin protein.12 Therefore, the creation of a favorable environment within the trauma is the first step in the promotion of wound healing, and treatment of infection is still key to trauma treatment.13
Clinically, the most common opportunistic infection pathogenic bacteria in hospital may be Pseudomonas aeruginosa, which is the main reason for the infection of the immunocompromised patients in hospitals. It is also the main cause of the prolonged unhealed infection of chronic decubitus ulcers and diabetic foot ulcers.2
During the early phase of trauma, neutrophils exude from the wound. It has been presumed that the major roles of neutrophils in wound repair are phagocytosis and bactericidal action. However, recent studies showed that the neutrophils also secrete tumor necrosis factor-α, interleukin-8, monocyte chemotactic protein-1, and other cytokines or bioactive substances.14Pseudomonas aeruginosa accelerated the formation of the biofilm to resist the monitoring of neutrophils, thereby escaping from the surveillance of antibiotics. The results of the current study showed that neutrophils increased sharply in both the control and infection groups, but increased more significantly in the infection group, which confirmed the phagocytosis and bactericidal action of neutrophils in the wound healing process.
The repair of damaged tissue is a very complex process, involving the interaction between the cells and the ECM. The wound infected by Pseudomonas aeruginosa in this study showed that on the first postoperative day, there was more purulent exudate in the wound, which is a sign of infection. The expression of TGF-β1 in the infected group was low in the first 3 days but then increased quickly and exceeded that of the control group at 7 days postoperation, indicating that the expression of TGF-β1 in the infected group was delayed. Moreover, the fact that the epithelization level was significantly lower than that of the control may associate closely with the delayed expression of TGF-β1. The role of changes of the extracellular environment on the epithelialization process cannot be ruled out, such as the inhibitory role of biofilm formation on epithelialization. The exact mechanism would be confirmed by a follow-up experiment.
In the process of wound healing, fibroblasts are activated to synthesize and secrete collagen. Transforming growth factor-β antagonizes the effects of inflammatory cytokines by inhibiting the expression of MMPs. It plays an important role in the ECM metabolism in the wound healing process and promotes wound healing and collagen deposition.15 Metalloproteinases are endopeptidases that utilize a Zn2+ or Ca2+ ion in their active site. These include the metzincin family of enzymes comprised of the serralysins, astacins, adamalysins (ie, a disintegrin and metalloproteinase), and matrixins (ie, MMPs).16 Matrix metalloproteinases are lowly expressed in the intact skin, but highly expressed if the skin was damaged by external stimuli.17 As the MMP becomes overexpressed, the rate of collagen degradation exceeds its synthesis and causes the formation of chronic skin ulcers.18,19 It was reported the occurrence of diabetic ulcers is correlated with the high expression of MMP-9,20 and the MMP-1 expression is 65 times higher in a diabetic ulcer than in a traumatic wound, but the TIMP-2 expression was reduced in the diabetic ulcer comparing to the traumatic wound.21 Overexpression and activation of MMPs and reduction of TIMPs lead to wound healing failure.22 Interleukin-8, granulocyte-macrophage colony-stimulating factor, and MMP-9 would release from the wound if it were infected with Pseudomonas aeruginosa or stimulated by cytokine.23 In the normal process of wound healing, MMP-2 promotes the remodeling of the ECM. The expression of MMP-2 can promote platelet aggregation and lead the scar tissue to transform into normal tissue.24-26 Matrix metalloproteinases-9, or gelatinase B, has been associated with wound repair and also been implicated in reepithelialization after injury.27,28
Conclusions
This study investigated the relationship between the formation of chronic Pseudomonas aeruginosa-infected refractory cutaneous wounds and TGF-β1 or MMPs. The authors found that TGF-β1 expression in the control group was much higher than the bacteria-infected group, which was in line with the previous findings that TGF-β1 expression was low in chronic wounds.29,30 Only a few basal cells in the normal skin epidermis layer of the posttraumatic and bacterial infection expressed MMP-9. Matrix metalloproteinase-9 was weakly expressed in the control dermal layer of the hair follicle, sebaceous glands, and vascular endothelial cells, but strongly expressed in the infected group. In the case of a weak inflammatory response of the control group, MMP-9 expression tends to be weak, and the ECM metabolism quickly reaches a state of equilibrium. While in the bacteria-infected group, MMP-9 expression increased and the wound healing delay may be related to the overexpression of MMP-9. This means the degradation rate was greater than the rate of synthesis, and therefore delayed the healing of wounds. In the control group, MMP-2 expression was lower than in the infected group, and the low expression of TGF-β1was related to the overexpression of MMPs. However, in the infected group, the low expression of TIMP-1 reduced the consumption of some MMPs, and also increased collagen degradation, delaying wound healing speed.
Extracellular matrix has both structural and regulatory roles, and has been emerging as a major research area in diverse aspects of ECM biology relevant to inflammation, tissue destruction, fibrosis, and regeneration.31 Tranforming growth factor-β has a complex role in fibrosis, inflammation, and cellular metaplasia. It is tethered to the ECM via latent TGF-β-binding proteins, which are anchored to fibrillin microfibrils.32 The experiments in the current study suggested that the wounded skin tissue infected with Pseudomonas aeruginosa delayed the expression of TGF-β1, and then caused the biology of the ECM to change relevant to fibrosis and regeneration, thus affecting the wound healing process.
Acknowledgments
Affiliation: The authors are from the China-Japan Union Hospital of Jilin University, Jilin, China.
Address correspondence to:
Guang Zhang, PhD
Department of Plastic and Reconstructive Surgery
China-Japan Union Hospital of Jilin University
No. 126 Xiantai Street
Changchun 130033
China
guangzhang@163.com
Disclosure: This study was supported by the National Natural Science Foundation of China (grant no. 81372068) and the International Cooperation Project by the Science and Technology Department of Jilin Province, China (no. 20130413013GH).
References
Sign Up Today





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
